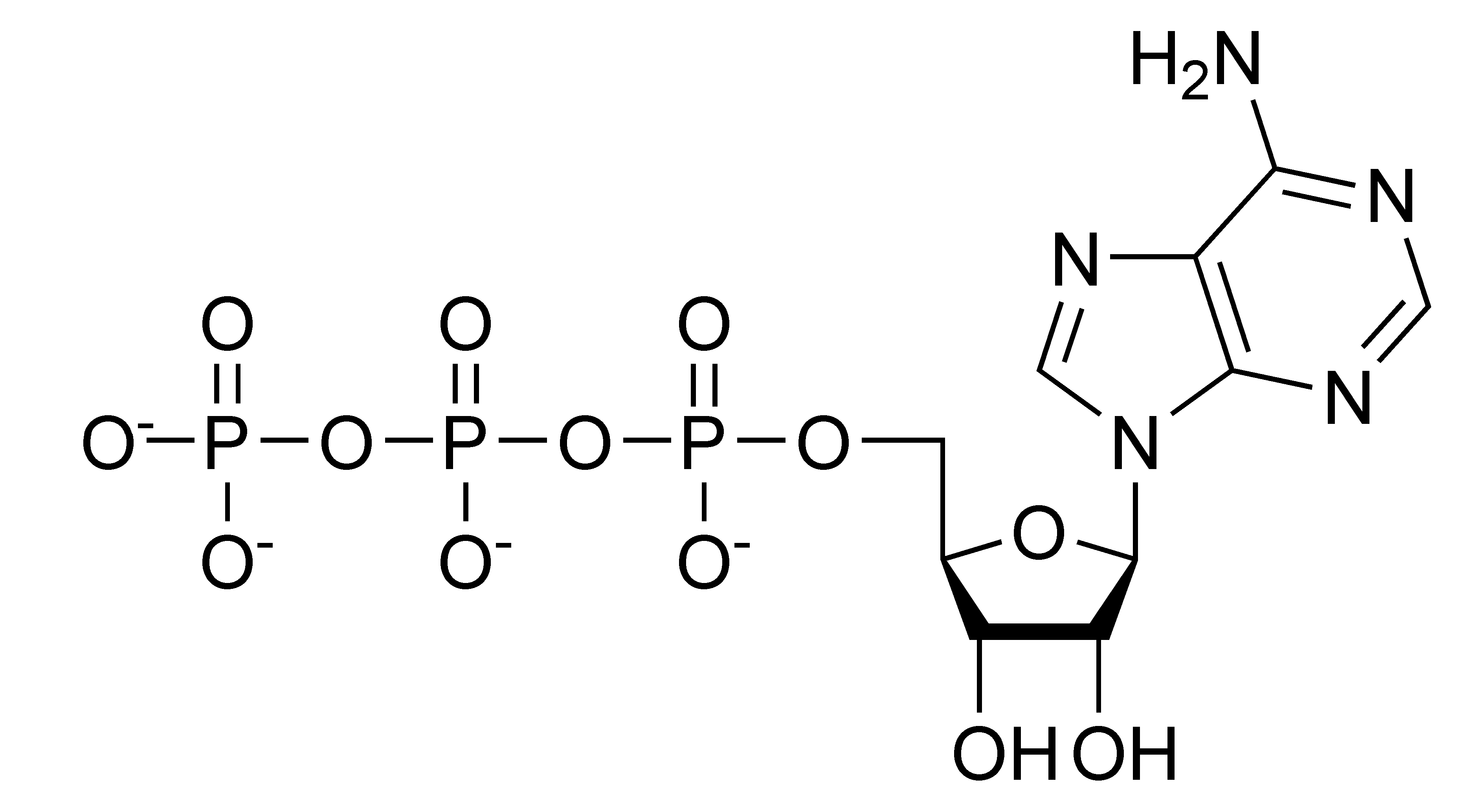
 In defining Biochemistry (previous Blog), one of the most important research questions that occupied the minds of the pioneers of the subject was how organisms extract energy from foods, such as carbohydrates, fats and proteins. Through a series of elegant experiments that occupied Biochemists during the first half of the 20th century, it became clear that complex carbohydrates (for example) are broken down through a series of enzyme catalysed steps, into a series of intermediates, leading ultimately to the production of ATP (top LHS), the universal energy currency in most living organisms (I say most because there are always exceptions!). I like to think of ATP generation more as a recycling reaction, in keeping with the theme of the First Law of Thermodynamics; which describes the conservation of energy in the Universe. Understanding the properties of ATP and similar biological nucleotides is an important aspect of Biochemistry and you can find a nice description of the chemistry and nomenclature of nucleotides at this link, provided by Dr. Scott Bellos an MD and former Chemistry Graduate at Rensselaer Polytechnic Institute in New York, whose learning resources in Nucleotide Biochemistry are highly recommended.
In defining Biochemistry (previous Blog), one of the most important research questions that occupied the minds of the pioneers of the subject was how organisms extract energy from foods, such as carbohydrates, fats and proteins. Through a series of elegant experiments that occupied Biochemists during the first half of the 20th century, it became clear that complex carbohydrates (for example) are broken down through a series of enzyme catalysed steps, into a series of intermediates, leading ultimately to the production of ATP (top LHS), the universal energy currency in most living organisms (I say most because there are always exceptions!). I like to think of ATP generation more as a recycling reaction, in keeping with the theme of the First Law of Thermodynamics; which describes the conservation of energy in the Universe. Understanding the properties of ATP and similar biological nucleotides is an important aspect of Biochemistry and you can find a nice description of the chemistry and nomenclature of nucleotides at this link, provided by Dr. Scott Bellos an MD and former Chemistry Graduate at Rensselaer Polytechnic Institute in New York, whose learning resources in Nucleotide Biochemistry are highly recommended.
The question I always ask myself, when I read about the
central role played by ATP in biological energy transactions, is why not GTP,
or CTP etc. The first point to consider is that the release of energy as the
phosphate ester bond is cleaved generates approximately 30kJ/mol of energy
(usually expressed as ΔGo' = -30.5 kJ/mol for ATP
hydrolysis to ADP and Pi). [I
shall comment on units in a separate studentmicroblog]. Is this the same for
GTP etc? The answer is of course yes, so there is nothing special about this particular nucleotide triphosphate. The preference for ATP in Biology, is probably just a chance, evolutionary event, in which adenine was fixed as the primary component of many biological cofactors and metabolites, including NAD, NADP, FAD, acetyl CoA etc. [Look them up on Google Images and make sure you can see the adenine moiety]. The evolution of adenine binding pockets in proteins was an important, early discovery in the development of X-ray Crystallography of proteins and has reinforced the priority position of adenine, over guanine etc in Biochemistry. The work of Michael Rossmann and John Walker, among others, was important in understanding the elements of primary and tertiary structure in protein recognition of adenine and its various derivatives (see image top RHS).
There are several factors that help us understand why the use of molecules like ATP has evolved to provide the fuel for many of the enzymatic steps that underpin metabolism.
1. ATP can be hydrolysed to release phosphate (Pi) yielding approximately 30kJ/mol of energy that can in turn be coupled to otherwise unfavourable reactions. In fact, it is not uncommon to find two hydrolysis steps (ATP to AMP) contributing 45kJ/mol more energy to an even more challenging biochemical reaction (the example that springs to my mind is the attachment of amino acids to their cognate transfer RNAs in protein biosynthesis). The work of Fritz Lipmann is of central importance in our appreciation of ATP in Bioenergetics. The ATP entry in Wikipedia is well worth an hour of your time on a Sunday morning!
2. The release of energy from the above hydrolysis reaction is a consequence of the relative "ease" with which ATP breaks down to ADP in water. It is important to realise that there is no "magical" packet of energy in ATP, but rather, in the aqueous, cellular environment at a pH slightly above neutrality, the equilibrium position of ATP=ADP +Pi, lies in favour of ADP, since the favourable interactions between water (Hydrogen bonds etc) and ADP are more stable than the covalently bonded phosphate. The repulsion between the phosphates in ATP, which is partly stabilised by Mg2+ ions, is also less entropically favoured over the hydrolysed products (ADP + Pi). The most important point to note is that ATP hydrolysis liberates sufficient energy to drive most of the thermodynamically unfavourable reactions in Biology. Occasionally ATP is hydrolysed to AMP to provide a "double shot" of energy and sometimes, special measures must be employed (which you will discover at University level). The evolution of metabolism has therefore been considerably constrained by the physical chemical properties of ATP and this is why it is such an important molecule for we Biochemists to understand. As an exercise, make a list of 10 reactions that incorporate ATP in living organisms.
 3. Think of ATP as a rechargeable battery. At any one time, each individual probably "carries" 25g of ATP in their cells. However, it has been shown that each of us recycles our own body weight in ATP every day! We now need to consider how the food we eat generates ATP and where in the cell this takes place. A point made clear by a landmark publication in the late 1950s by Hans Krebs and Hans Kornberg (whose names you will find on the Innovation Lab bench and who [a few years before the above picture was taken) used to be the head of the Sheffield students union!), where they set out very elegantly, the underlying principles of energy metabolism in living cells.
3. Think of ATP as a rechargeable battery. At any one time, each individual probably "carries" 25g of ATP in their cells. However, it has been shown that each of us recycles our own body weight in ATP every day! We now need to consider how the food we eat generates ATP and where in the cell this takes place. A point made clear by a landmark publication in the late 1950s by Hans Krebs and Hans Kornberg (whose names you will find on the Innovation Lab bench and who [a few years before the above picture was taken) used to be the head of the Sheffield students union!), where they set out very elegantly, the underlying principles of energy metabolism in living cells.  We shall use the classic "Boehringer Mannheim" Metabolic Map, first produced by Gerhard Michal in 1965 (so we are celebrating its 50th Birthday this year!), which I think came out of the earlier Nicholson maps in the late 1950s, to illustrate the networks and inter-relationships between metabolic pathways. Think of metabolism as the London Underground, or the Paris Metro. The important points about metabolic pathways is that they are rarely free-standing and they comprise a series of enzyme catalysed reactions, some of which are regulated. [The mechanisms of regulation are diverse and can include chemical modification of the enzyme, controlled hydrolysis or synthesis to regulate the "amount" of enzyme [I shall discuss steady state phenomena in a separate Blog]. It was also discovered in the 1960s that some enzymes can be "modulated" by small molecules that are distinct from the substrates or products: we refer to these as allosteric effectors and they can either activate or inhibit an enzyme catalysed reaction]. Importantly, the conversion of molecules like complex carbohydrates takes place via a series of reactions, some of which decompose and some modify the intermediates. The purpose of these pathways is to generate energy (ATP) or to recycle the carbohydrate into the molecules that make up the organism. This includes other metabolites, other carbohydrates, fats, proteins etc. This is why an appreciation of metabolism is such a defining part of Biochemistry. Moreover, the malfunctioning of metabolism and its regulation lie at the heart of many diseases and is critically important in drug discovery, which will be discussed to illuminate several aspects of metabolic principles.
We shall use the classic "Boehringer Mannheim" Metabolic Map, first produced by Gerhard Michal in 1965 (so we are celebrating its 50th Birthday this year!), which I think came out of the earlier Nicholson maps in the late 1950s, to illustrate the networks and inter-relationships between metabolic pathways. Think of metabolism as the London Underground, or the Paris Metro. The important points about metabolic pathways is that they are rarely free-standing and they comprise a series of enzyme catalysed reactions, some of which are regulated. [The mechanisms of regulation are diverse and can include chemical modification of the enzyme, controlled hydrolysis or synthesis to regulate the "amount" of enzyme [I shall discuss steady state phenomena in a separate Blog]. It was also discovered in the 1960s that some enzymes can be "modulated" by small molecules that are distinct from the substrates or products: we refer to these as allosteric effectors and they can either activate or inhibit an enzyme catalysed reaction]. Importantly, the conversion of molecules like complex carbohydrates takes place via a series of reactions, some of which decompose and some modify the intermediates. The purpose of these pathways is to generate energy (ATP) or to recycle the carbohydrate into the molecules that make up the organism. This includes other metabolites, other carbohydrates, fats, proteins etc. This is why an appreciation of metabolism is such a defining part of Biochemistry. Moreover, the malfunctioning of metabolism and its regulation lie at the heart of many diseases and is critically important in drug discovery, which will be discussed to illuminate several aspects of metabolic principles.
I shall discuss enzyme catalysis in the next post.
Key points
Energy in living organisms is stored and released in the form of ATP
The hydrolysis of ATP is harnessed to unfavourable biochemical reactions (this is called coupling)
Foods like carbohydrates and fats are "processed" by all living organisms by a set of enzyme catalysed reactions to generate NADH and FADH, both of which provide the route to ATP synthesis on the inner mitochondrial membrane.
These pathways are interconnected, complex and serve not only to produce energy, but to build the organism, both structurally (skin and bones, or leaves and stems) and functionally (enzymes and genes).
The metabolic pathways that were charted in the last century are generally found in all living organisms (aerobic and anaerobic organisms do have significant differences), but there are some interesting and instructive exceptions.
The regulation of metabolism by a range of Biochemical strategies is an important phenomenon that has become a key aspect of drug discovery since it is often faulty in a number of diseases.
http://lutcbtec.blogspot.com/2015/01/the-biochemical-basis-of-energy.html
ReplyDelete